- это сложные мономеры, из которых собраны гетерополимерные молекулы. ДНК и РНК. Свободные нуклеотиды участвуют в сигнальных и энергетических процессах жизнедеятельности. ДНК-нуклеотиды и РНК-нуклеотиды имеют общий план строения, но различаются по строению сахара-пентозы. В ДНК-нуклеотидах используется сахар дезоксирибоза, а в РНК-нуклеотидах - рибоза.
Структура нуклеотида
В каждом нуклеотиде можно выделить 3 части:
1. Углевод - это пятичленный сахар-пентоза (рибоза или дезоксирибоза).
2. Фосфорный остаток (фосфат) - это остаток фосфорной кислоты.
3. Азотистое основание - это соединение, в котором много атомов азота. В нуклеиновых кислотах используется всего 5 видов азотистых оснований: Аденин, Тимин, Гуанин, Цитозин, Урацил. В ДНК - 4 вида: Аденин, Тимин, Гуанин, Цитозин. В РНК - тоже 4 вида: Аденин, Урацил, Гуанин, Цитозин, Легко заметить, что в РНК происходит замещение Тимина на Урацил по сравнению с ДНК.
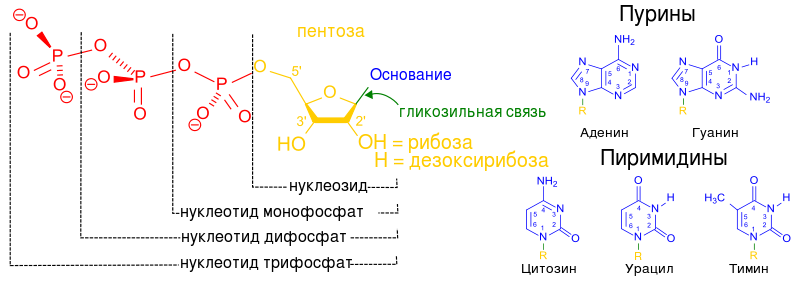
Общая структурная формула пентозы (рибозы или дезоксирибозы), молекулы которой образуют "скелет" нуклеиновых кислот:
Если Х заменить на Н (Х = Н) - то получаются дезоксирибонуклеозиды; если Х заменить на ОН (Х = ОН) - то получаются рибонуклеозиды. Если вместо R подставить азотистое основание (пуриновое или пиримидиновое) - то получится конкретный нуклеотид.
Важно обратить внимание на те положения атомов углерода в пентозе, которые обозначены как 3" и 5". Нумерация атомов углерода начинается от атома кислорода вверху и идёт по часовой стрелке. Последним получается атом углерода (5"), который располагается за пределами пентозного кольца и образует, можно сказать, "хвостик" у пентозы. Так вот, при наращивании цепочки из нуклеотидов фермент может присоединить новый нуклеотид только к углероду 3" и ни к какому другому. Поэтому 5"-конец нуклеотидной цепочки никогда не сможет иметь продолжения, удлинняться может только 3"-конец.


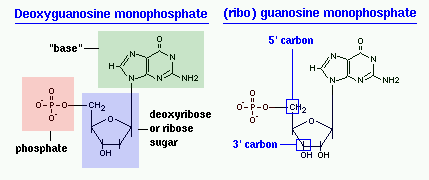
Сравните нуклеотид для РНК с нуклеотидом для ДНК.
Попробуйте узнать, какой это нуклеотид, в таком представлении:

АТФ - свободный нуклеотид
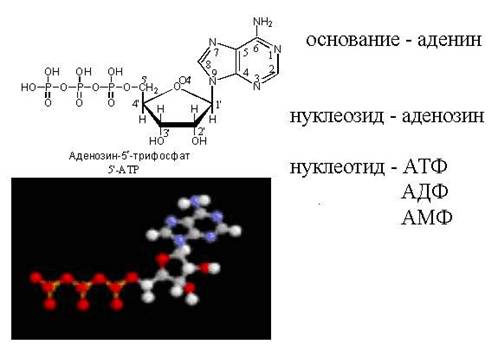
цАМФ - "закольцованная" молекула АТФ

Схема строения нуклеотида
Обратите внимание на то, что активированный нуклеотид, способный наращивать цепочку ДНК или РНК, имеет "трифосфатный хвостик". Именно этим "энергонасыщенным" хвостиком он может присоединиться к уже имеющейся цепочке растущей нуклеиновой кислоты. Фосфатный хвостик сидит на 5-м атоме углерода, так что это положение углерода уже занято фосфатами и предназнено для прикрепления. К чему же его прикрепить? Только к углероду в положении 3". После прикрепления данный нуклеотид сам станет мишенью дла прикрепления следующего нуклеотида. "Принимающая сторона" предоставляет углерод в положении 3", а "прибывающая сторона" цепляется к нему фосфатным хвостиком, находящимся в положении 5". В целом цепочка растёт со стороны 3".
Наращивание нуклеотидной цепочки ДНК
Наращивание цепочки за счёт "продольных" связей между нуклеотидами может идти только в одном направлении: от 5" ⇒ к 3", т.к. новый нуклеотид можно присоединить только к 3"-концу цепочки, но не к 5"-концу.
Пары нуклеотидов, связанные "поперечными" комплементарными связями своих азотистых оснований

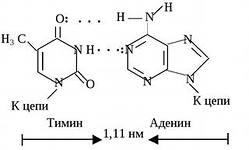
Участок двойной спирали ДНК
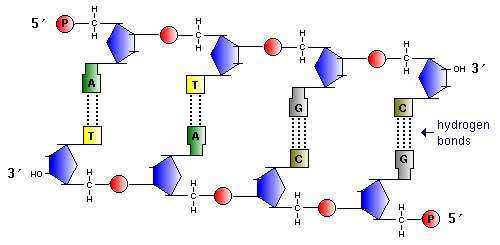
Найдите признаки антипараллельности двух цепей ДНК.
Найдите пары нуклеотидов с двойными и тройными комплементарными связями.
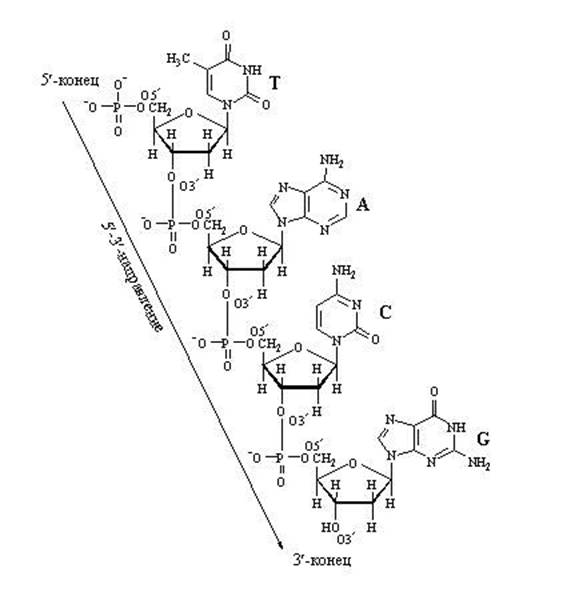
4.2.1. Первичной структурой нуклеиновых кислот называется последовательность расположения мононуклеотидов в цепи ДНК или РНК . Первичная структура нуклеиновых кислот стабилизируется 3",5"-фосфодиэфирными связями. Эти связи образуются при взаимодействии гидроксильной группы в 3"-положении пентозного остатка каждого нуклеотида с фосфатной группой соседнего нуклеотида (рисунок 3.2),
Таким образом, на одном конце полинуклеотидной цепи имеется свободная 5"-фосфатная группа (5"-конец), а на другом - свободная гидроксильная группа в 3"-положении (3"-конец). Нуклеотидные последовательности принято записывать в направлении от 5"-конца к 3"-концу.
Рисунок 4.2. Структура динуклеотида, в состав которого входят аденозин-5"-монофосфат и цитидин-5"-монофосфат.
4.2.2. ДНК (дезоксирибонуклеиновая кислота) содержится в клеточном ядре и имеет молекулярную массу порядка 1011 Да. В состав её нуклеотидов входят азотистые основания аденин, гуанин, цитозин, тимин , углевод дезоксирибоза и остатки фосфорной кислоты. Содержание азотистых оснований в молекуле ДНК определяют правила Чаргаффа:
1) количество пуриновых оснований равно количеству пиримидиновых (А + Г = Ц + Т) ;
2) количество аденина и цитозина равно количеству тимина и гуанина соответственно (А = Т; Ц = Г) ;
3) ДНК, выделенные из клеток различных биологических видов, отличаются друг от друга величиной коэффициента специфичности:
(Г + Ц) /(А + Т)
Эти закономерности в строении ДНК объясняются следующими особенностями её вторичной структуры:
1) молекула ДНК построена из двух полинуклеотидных цепей, связанных между собой водородными связями и ориентированных антипараллельно (то есть 3"-конец одной цепи расположен напротив 5"-конца другой цепи и наоборот);
2) водородные связи образуются между комплементарными парами азотистых оснований. Аденину комплементарен тимин; эта пара стабилизируется двумя водородными связями. Гуанину комплементарен цитозин; эта пара стабилизируется тремя водородными связями (см. рисунок б) . Чем больше в молекуле ДНК пар Г-Ц, тем больше её устойчивость к действию высоких температур и ионизирующего излучения;

Рисунок 3.3. Водородные связи между комплементарными азотистыми основаниями.
3) обе цепи ДНК закручены в спираль, имеющую общую ось. Азотистые основания обращены внутрь спирали; кроме водородных, между ними возникают также гидрофобные взаимодействия. Рибозофосфатные части расположены по периферии, образуя остов спирали (см. рисунок 3.4).

Рисунок 3.4. Схема строения ДНК.
4.2.3. РНК (рибонуклеиновая кислота) содержится преимущественно в цитоплазме клетки и имеет молекулярную массу в пределах 104 - 106 Да. В состав её нуклеотидов входят азотистые основания аденин, гуанин, цитозин, урацил , углевод рибоза и остатки фосфорной кислоты. В отличие от ДНК, молекулы РНК построены из одной полинуклеотидной цепи, в которой могут находиться комплементарные друг другу участки (рисунок 3.5). Эти участки могут взаимодействовать между собой, образуя двойные спирали, чередующиеся с неспирализованными участками.

Рисунок 3.5. Схема строения транспортной РНК.
По особенностям структуры и функции различают три основных типа РНК:
1) матричные (информационные) РНК (мРНК) передают информацию о структуре белка из клеточного ядра на рибосомы;
2) транспортные РНК (тРНК) осуществляют транспорт аминокислот к месту синтеза белка;
3) рибосомальные РНК (рРНК) входят в состав рибосом, участвуют в синтезе белка.
Нуклеиновые кислоты представляют собой высокомолекулярные соединения, молекулярная масса которых колеблется от 25 тыс. до 1 млн и более.
Полимерные цепи нуклеиновых кислот построены из мономерных единиц - нуклеотидов, в связи с чем нуклеиновые кислоты назы- вают полинуклеотидами.
Обычно «неделимое» мономерное звено (например, аминокислотный остаток в белках) у нуклеотидов представляет собой трехкомпонентное образование, включающее гетероциклическое основание, углеводный остаток и фосфатную группу.
Углеводными компонентами служат пентозы - D-рибоза и 2-дезокси-э-рибоза. В зависимости от этого нуклеиновые кислоты делятся на рибонуклеиновые (РНК), содержащие рибозу, и дезоксирибо- нуклеиновые (ДНК), содержащие дезоксирибозу.
ДНК содержатся в основном в ядрах клеток, РНК находятся преимущественно в рибосомах, а также протоплазме клеток. РНК непосредственно участвуют в биосинтезе белка.
14.1. Нуклеотиды
14.1.1. Нуклеозиды
В химии нуклеиновых кислот входящие в их состав гетероциклические соединения пиримидинового и пуринового рядов обычно называют нуклеиновыми основаниями.
Нуклеиновые основания в качестве заместителей в гетероцикле могут содержать:
Либо оксогруппу, как в урациле и тимине;
Либо аминогруппу, как в аденине;
Либо одновременно обе эти группы, как в цитозине и гуанине.
Кислородсодержащие основания представлены лактамными таутомерными формами, в которых ароматичность не нарушена (см. 13.4). Для всех оснований приняты сокращенные трехбуквенные обозначения, составленные из первых букв их латинских названий.
Нуклеиновые кислоты различаются входящими в них гетероциклическими основаниями: урацил входит только в РНК, а тимин -
в ДНК:

Нуклеиновые основания образуют связь за счет одного из атомов азота с аномерным центром пентозы (D-рибозы или 2-дезокси-D- рибозы). Этот тип связи аналогичен обычной гликозидной связи и известен как N-гликозидная связь, а сами гликозиды - как N-гликози- ды. В химии нуклеиновых кислот их называют нуклеозидами.
В состав природных нуклеозидов пентозы входят в фуранозной форме (атомы углерода в них нумеруют цифрой со штрихом). Гликозидная связь осуществляется с атомом азота N-1 пиримидинового и N-9 пуринового оснований.

Природные нуклеозиды всегда являются β -аномерами.
В зависимости от природы углеводного остатка различают рибонуклеозиды и дезоксирибонуклеозиды. Для нуклеозидов употребительны названия, производимые от тривиального названия соответствующего нуклеинового основания с суффиксами -идин у пиримидиновых и -озин у пуриновых нуклеозидов.

Исключение составляет название «тимидин» (а не дезокситимидин), используемое для дезоксирибозида тимина, входящего в состав ДНК. В тех редких случаях, когда тимин встречается в РНК, соответствующий нуклеозид называется риботимидином.
Трехбуквенные символы нуклеозидов отличаются от символов оснований последней буквой. Однобуквенные символы применяются только для остатков (радикалов) нуклеозидов в более сложных структурах.
Нуклеозиды устойчивы к гидролизу в слабощелочной среде, но гидролизуются в кислой. Пуриновые нуклеозиды гидролизуются легко, пиримидиновые труднее.
В качестве лекарственных средств в онкологии используют синтетические производные пиримидинового и пуринового рядов, по строению похожие на естественные метаболиты (в данном случае - на нуклеиновые основания), но не полностью им идентичные, т. е. являющиеся антиметаболитами. Например, 5-фторурацил выступает

в роли антагониста урацила и тимина, 6-меркаптопурин - аденина. Конкурируя с метаболитами, они нарушают синтез нуклеиновых кислот в организме на разных этапах.

14.1.2. Нуклеотиды
Нуклеотидами называют фосфаты нуклеозидов. Фосфорная кислота обычно этерифицирует спиртовый гидроксил при С-5" или С-3" в остатке рибозы (рибонуклеотиды) или дезоксирибозы (дезоксирибонуклеотиды).
Общий принцип строения нуклеотидов показан на примере фосфатов аденозина. Для связывания трех компонентов в молекуле нуклеотида используются сложноэфирная и N-гликозидная связи.
Нуклеотиды можно рассматривать, с одной стороны, как эфиры нуклеозидов (фосфаты), а с другой - как кислоты (в связи с наличием остатка фосфорной кислоты).

За счет фосфатного остатка нуклеотиды проявляют свойства двухосновной кислоты и в физиологических условиях при рН ~7 находятся в полностью ионизированном состоянии.

Для нуклеотидов используют два вида названий (табл. 14.1). Одно включает наименование нуклеозида с указанием положения в нем фосфатного остатка, например, аденозин-3"-фосфат, уридин-5"-фос- фат; другое строится с добавлением сочетания -иловая кислота к названию остатка пиримидинового основания, например, 5"-уридило- вая кислота, или пуринового основания, например 3"-адениловая кислота.
Используя принятый для нуклеозидов однобуквенный код, 5"-фосфаты записывают с добавлением латинской буквы «р» перед символом нуклеозида, 3"-фосфаты - после символа нуклеозида. Аденозин-5"-фосфат обозначается рА, аденозин-3"-фосфат - Ар и т. п. Эти сокращенные обозначения используют для записи последовательности нуклеотидных остатков в нуклеиновых кислотах. По отношению к свободным нуклеотидам в биохимической литера-
туре широко используют их названия как монофосфатов с отражением этого признака в сокращенном коде, например АМР (или АМФ) для аденозин-5"-фосфата и т. д. (см. табл. 14.1).
Таблица 14.1. Важнейшие нуклеотиды, входящие в состав нуклеиновых кислот

Циклофосфаты. К ним относятся нуклеотиды, у которых одна молекула фосфорной кислоты этерифицирует одновременно две гидроксильные группы углеводного остатка. Практически во всех клетках присутствуют два нуклеозидциклофосфата - аденозин-3",5"- циклофосфат (cAMP) и гуанозин-3",5"-циклофосфат (cGMP).

14.2. Структура нуклеиновых кислот
14.2.1. Первичная структура
В полинуклеотидных цепях нуклеотидные звенья связаны через фосфатную группу. Фосфатная группа образует две сложноэфирные связи: с С-3" предыдущего и с С-5" последующего нуклеотидных звеньев (рис. 14.1). Каркас цепи состоит из чередующихся пентозных и фосфатных остатков, а гетероциклические основания являются «боковыми» группами, присоединенными к пентозным остаткам. Нуклеотид со свободной 5"-ОН группой называют 5"-концевым, а нуклеотид со свободной З"-ОН группой - З"-концевым.
На рисунке 14.2 приведено строение произвольного участка цепи ДНК, включающего четыре нуклеиновых основания. Легко представить, какое множество сочетаний можно получить путем варьирования последовательности четырех нуклеотидных остатков. Принцип построения цепи РНК такой же, как и у ДНК, с двумя исключениями: пентозным остатком в РНК служит D-рибоза, а в наборе гетероциклических оснований используется не тимин, а урацил.
Первичная структура нуклеиновых кислот определяется последовательностью нуклеотидных звеньев, связанных ковалентными связями в непрерывную цепь полинуклеотида.
Для удобства записи первичной структуры существует несколько способов сокращений. Один из них заключается в использовании ранее приведенных сокращенных названий нуклеозидов. Например, показанный на рис. 14.2 фрагмент цепи ДНК может быть записан

Рис. 14.1. Общий принцип строения полинуклеотидной цепи

Рис. 14.2. Первичная структура участка цепи ДНК
как d(ApCpGpTp...) или d(A-C-G-T...). Часто букву d опускают, если очевидно, что речь идет о ДНК.
Важной характеристикой нуклеиновых кислот служит нуклеотидный состав, т. е. набор и количественное отношение нуклеотидных компонентов. Нуклеотидный состав устанавливают, как правило, путем исследования продуктов гидролитического расщепления нуклеиновых кислот.
ДНК и РНК различаются поведением в условиях щелочного и кислотного гидролиза. ДНК устойчивы к гидролизу в щелочной среде. РНК легко гидролизуются в мягких условиях в щелочной среде до нуклеотидов, которые, в свою очередь, способны в щелочной среде отщеплять остаток фосфорной кислоты с образованием нуклеозидов. Нуклеозиды в кислой среде гидролизуются до гетероциклических оснований и углеводов.

14.2.2. Вторичная структура ДНК
Под вторичной структурой понимают пространственную организацию полинуклеотидной цепи. Согласно модели Уотсона-Крика молекула ДНК состоит из двух полинуклеотидных цепей, правозакрученных вокруг общей оси с образованием двойной спирали. Пуриновые и пиримидиновые основания направлены внутрь спирали. Между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи возникают водородные связи. Эти основа- ния составляют комплементарные пары.
Водородные связи образуются между аминогруппой одного основания и карбонильной группой другого -NH...O=C- , а также между амидным и иминным атомами азота -NH...N-Например, как показано ниже, между аденином и тимином образуются две водородные связи, и эти основания составляют комплементарную пару, т. е. аденину в одной цепи будет соответствовать тимин в другой цепи. Другую пару комплементарных оснований составляют гуанин и цитозин, между которыми возникают три водородные связи.

Водородные связи между комплементарными основаниями - один из видов взаимодействий, стабилизирующих двойную спираль. Две цепи ДНК, образующие двойную спираль, не идентичны, но комплементарны между собой. Это означает, что первичная структура, т. е. нуклеотидная последовательность, одной цепи предопределяет первичную структуру второй цепи (рис. 14.3).

Рис. 14.3. Комплементарность полинуклеотидных цепей в двойной спирали
ДНК
14.3. Нуклеотидные коферменты
Нуклеотиды имеют большое значение не только как строительный материал для нуклеиновых кислот. Они участвуют в биохими- ческих процессах и особенно важны в роли коферментов, т. е. веществ, тесно связанных с ферментами и необходимых для проявления ими ферментативной активности.
14.3.1. Нуклеозидполифосфаты
Во всех тканях организма содержатся моно-, ди- и трифосфаты нуклеозидов. Особенно широко известны аденинсодержащие нук- леотиды - аденозин-5"-фосфат (АМР), аденозин-5"-дифосфат (ADP)
и аденозин-5"-трифосфат (ATP) (для этих соединений наряду с приведенными сокращенными обозначениями латинскими буквами в оте- чественной литературе используют сокращения соответствующих русских названий - АМФ, АДФ, АТФ).
Нуклеотиды, фосфорилированные в разной степени, способны к взаимопревращениям путем наращивания или отщепления фос- фатных групп. Дифосфатная группа содержит одну, а трифосфатная - две ангидридные связи, называемые макроэргическими, поскольку они обладают большим запасом энергии. Необходимые для образования такой связи энергетические затраты восполняются за счет энергии, выделяемой в процессе метаболизма углеводов. При расщеплении макроэргической связи Р~О (обозначаемой волнистой линией) выделяется ~32 кДж/моль. С этим связана важнейшая роль АТФ как «поставщика» энергии во всех живых клетках.
В показанных ниже взаимопревращениях АМФ, АДФ и АТФ формулы этих соединений соответствуют их неионизированному состоянию. В физиологических условиях при рН ~7 фосфатные группы почти полностью ионизированы, поэтому в биохимической литературе эти и любые другие нуклеотиды записывают соответственно в виде анионов.

Нуклеозидполифосфаты в биохимических процессах. С участием АТФ и АДФ в организме осуществляется важнейший биохимический процесс - перенос фосфатных групп. Например, образование сложных эфиров (фосфатов) - типичная реакция в метаболизме углеводов. Все стадии гликолиза (превращения глюкозы в пируват) осуществляются только в фосфатной форме. Получение фосфатов гидроксилсодержа- щих соединений можно представить в виде общей схемы.
Так, галактоза, образующаяся при расщеплении лактозы, на начальной стадии метаболического превращения в глюкозу взаимо- действует с АТФ с образованием монофосфата.

14.3.2. Никотинамиднуклеотиды
Наиболее важными представителями этой группы соединений являются никотинамидадениндинуклеотид (NAD, или в русской литературе НАД) и его фосфат (NADP, или НАДФ). Эти соединения выполняют важную роль коферментов в осуществлении многих
окислительно-восстановительных реакций. В соответствии с этим они могут существовать как в окисленной (НАД+, НАДФ+), так и восстановленной (НАДН, НАДФН) форме.

Структурным фрагментом НАД + и НАДФ + является никотинамидный остаток в виде пиридиниевого катиона. В составе НАДН и НАДФН этот фрагмент превращается в остаток 1,4-дигидропиридина.
В ходе биологического дегидрирования субстрат теряет два атома водорода, т. е. два протона и два электрона (2Н+, 2е) или протон и гидрид-ион (Н+ и Н -). Кофермент НАД+ обычно рассматривается как акцептор гидрид-иона Н - (хотя окончательно не установлено, происходит ли перенос атома водорода к этому коферменту одновременно с переносом электрона или эти процессы протекают раздельно).

В результате восстановления путем присоединения гидрид-иона к НАД+ пиридиниевое кольцо переходит в 1,4-дигидропиридиновый фрагмент. Этот процесс обратим.

В реакции окисления ароматический пиридиниевый цикл переходит в неароматический 1,4-дигидропиридиновый цикл. В связи с потерей ароматичности возрастает энергия НАДН по сравнению с НАД + . Таким способом НАДН запасает энергию, которая затем расходуется в других биохимических процессах, требующих энергетических затрат.
Типичными примерами биохимических реакций с участием НАД+ служат окисление спиртовых групп в альдегидные (например, пре- вращение ретинола в ретиналь, см. 15.4), а с участием НАДН - восстановление карбонильных групп в спиртовые (превращение пировиноградной кислоты в молочную, см. 9.2.3).
Лекция № 19
НУКЛЕОЗИДЫ. НУКЛЕОТИДЫ. НУКЛЕИНОВЫЕ КИСЛОТЫ
План
- Нуклеиновые основания.
- Нуклеозиды.
- Нуклеотиды.
- Нуклеотидные коферменты.
- Нуклеиновые кислоты.
Лекция № 19
НУКЛЕОЗИДЫ. НУКЛЕОТИДЫ. НУКЛЕИНОВЫЕ
КИСЛОТЫ
План
- Нуклеиновые основания.
- Нуклеозиды.
- Нуклеотиды.
- Нуклеотидные коферменты.
- Нуклеиновые кислоты.
Нуклеиновые кислоты – присутствующие в
клетках всех живых организмов биополимеры, которые выполняют важнейшие функции
по хранению и передаче генетической информации и участвуют в механизмах ее
реализации в процессе синтеза клеточных белков.
Установление состава нуклеиновых кислот путем их последовательного
гидролитического расщепления позволяет выделить следующие структурные
компоненты.
Рассмотрим структурные компоненты нуклеиновых
кислот в порядке усложнения их строения.
1. Нуклеиновые основания.
Гетероциклические основания, входящие в состав
нуклеиновых кислот (нуклеиновые основания
), — это гидрокси- и
аминопроизводные пиримидина и пурина. Нуклеиновые кислоты содержат три
гетероциклических основания с пиримидиновым циклом (пиримидиновые
основания
) и два — с пуриновым циклом
(пуриновые основания
).
Нуклеиновые основания
имеют тривиальные названия и соответствующие однобуквенные обозначения.

В составе нуклеиновых кислот гетероциклические
основания находятся в термодинамически стабильной оксоформе.
Кроме этих групп нуклеиновых оснований,
называемых основными
, в нуклеиновых кислотах в небольших количествах
встречаются минорные
основания: 6-оксопурин (гипоксантин),
3-N-метилурацил, 1-N-метилгуанин и др.
Нуклеиновые кислоты включают остатки
моносахаридов – D-рибозы и 2-дезокси –D-рибозы. Оба моносахарида присутствуют в
нуклеиновых кислотах в
b
-фуранозной форме.

2. Нуклеозиды.
Нуклеозиды – это N-гликозиды, образованные нуклеиновыми основаниями и рибозой
или дезоксирибозой.
Между аномерным атомом углерода моносахарида и атомом азота в положении 1
пиримидинового цикла или атомом азота в положении 9 пуринового цикла образуется b
-гликозидная
связь.

В зависимости от природы моносахаридного остатка
нуклеозиды делят на рибонуклеозиды
(содержат остаток рибозы) и дезоксирибонуклеозиды
(содержат остаток дезоксирибозы). Названия
нуклеозидов строят на основе тривиальных названий нуклеиновых оснований,
добавляя окончание –идин
для производных пиримидина и -озин
для
производных пурина. К названиям дезоксирибонуклеозидов добавляют приставку дезокси-.
Исключение составляет нуклеозид, образованный тимином и
дезоксирибозой, к которому приставка дезокси-
не добавляется, так как
тимин образует нуклеозиды с рибозой лишь в очень редких случаях.

Для обозначения нуклеозидов используются
однобуквенные обозначения, входящих в их состав нуклеиновых оснований. К
обозначениям дезоксирибонуклеозидов (за исключением тимидина) добавляется буква
”д”.
Наряду с представленными на схеме основными
нуклеозидами в составе нуклеиновых кислот встречаются минорные нуклеозиды,
содержащие модифицированные нуклеиновые основания (см. выше).
В природе нуклеозиды встречаются также в
свободном состоянии, преимущественно в виде нуклеозидных антибиотиков, которые
проявляют противоопухолевую активность. Нуклеозиды-антибиотики имеют некоторые
отличия от обычных нуклеозидов в строении либо углеводной части, либо
гетероциклического основания, что позволяет им выступать в качестве
антиметаболитов, чем и объясняется их антибиотическая активность.
Как N-гликозиды, нуклеозиды устойчивы к действию
щелочей, но расщепляются под действием кислот с образованием свободного
моносахарида и нуклеинового основания. Пуриновые нуклеозиды гидролизуются
значительно легче пиримидиновых.
3. Нуклеотиды
Нуклеотиды – это эфиры нуклеозидов и фосфорной
кислоты (нуклеозидфосфаты). Сложноэфирную связь с фосфорной кислотой образует ОН
группа в положении 5
/ или
3
/ моносахарида. В зависимости от
природы моносахаридного остатка нуклеотиды делят на рибонуклеотиды
(структурные элементы РНК) и дезоксирибонуклеотиды
(структурные элементы
ДНК). Названия нуклеотидов включают название нуклеозида с указанием положения в
нем остатка фосфорной кислоты. Сокращенные обзначения нуклеозидов содержат
обозначение нуклеозида, остатка моно-, ди- или трифосфорной кислоты, для
3
/ -производных указывается также
положение фосфатной группы.

Нуклеотиды являются мономерными звеньями, из
которых построены полимерные цепи нуклеиновых кислот. Некоторые нуклеотиды
выполняют роль коферментов и участвуют в обмене веществ.
4. Нуклеотидные
коферменты
Коферменты
– это органические соединения
небелковой природы, которые необходимы для осуществления каталитического
действия ферментов. Коферменты относятся к разным классам органических
соединений. Важную группу коферментов составляют нуклеозидполифосфаты
.
Аденозинфосфаты –
производные
аденозина, содержащие остатки моно-, ди- и трифосфорных кислот. Особое место
занимают аденозин-5 / -моно-, ди- и
трифосфаты — АМФ, АДФ и АТФ — макроэргические
вещества, которые обладают
большими запасами свободной энергии в подвижной форме. Молекула АТФ содержит
макроэргические связи Р-О, которые легко расщепляются в результате гидролиза.
Выделяющаяся при этом свободная энергия обеспечивает протекание сопряженных с
гидролизом АТФ термодинамически невыгодных анаболических процессов, например,
биосинтез белка.

Кофермент А
.
Молекула этого
кофермента состоит из трех структурных компонентов: пантотеновой кислоты,
2-аминоэтантиола и АДФ.

Кофермент А участвует в процессах
ферментативного ацилирования, активируя карбоновые кислоты путем превращения их
в реакционноспособные сложные эфиры тиолов.
Никотинамидадениндинуклеотидные коферменты.
Никотинамидадениндинуклеотид (НАД +)
и его фосфат (НАДФ
+ ) содержат в своем составе катион пиридиния в виде
никотинамидного фрагмента. Пиридиниевый катион в составе этих коферментов
способен обратимо присоединять гидрид-анион с образованием восстановленной формы
кофермента — НАД
Н.

Таким образом никотинамидадениндинуклеотидные
коферменты участвуют в окислительно-восстановительных процессах, связанных с
переносом гидрид-аниона, например, окислении спиртовых групп в альдегидные
(превращение ретинола в ретиналь), восстановительном аминировании кетокислот,
восстановлении кетокислот в гидроксикислоты. В ходе этих процессов субстрат
теряет (окисление) или присоединяет (восстановление) два атома водорода в виде
Н
+ и Н
— . Кофермент служит при этом акцептором
(НАД
+ ) или донором
(НАД .
Н) гидрид-иона. Все процессы с
участием коферментов являются стереоселективными. Так, при восстановлении
пировиноградной кислоты образуется исключительно L-молочная кислота.

5. Нуклеиновые кислоты.
Первичная структура
нуклеиновых кислот представляет собой линейную полимерную цепь, построенную
из мономеров – нуклеотидов, которые связаны между собой
3 / -5 / -фосфодиэфирными
связями. Полинуклеотидная цепь имеет 5′-конец и 3′- конец. На 5′-конце находится
остаток фосфорной кислоты, а на 3′- конце — свободная гидроксильная группа.
Нуклеотидную цепь принято записывать, начиная с 5′-конца.

В зависимости от природы моносахаридных остатков
в нуклеотиде различают дезоксирибонуклеиновые кислоты (ДНК) и рибонуклеиновые
кислоты (РНК). ДНК и РНК различаются также по природе входящих в их состав
нуклеиновых оснований: урацил входит только в состав РНК, тимин – только в
состав ДНК.
Вторичная структура
ДНК
представляет собой комплекс двух полинуклеотидных цепей, закрученных вправо
вокруг общей оси так, что углевод-фосфатные цепи находятся снаружи, а
нуклеиновые основания направлены внутрь (двойная спираль Уотсона-Крика
).
Шаг спирали — 3.4 нм, на 1 виток приходится 10 пар нуклеотидов. Полинуклеотидные
цепи антипараллельны,т.е.
напротив 3′-конца одной цепи находится 5′-конец другой цепи. Две цепи ДНК
неодинаковы по своему составу, но они комплементарны
. Это выражается в
том, что напротив аденина (А) в одной цепи всегда находится тимин (Т) в другой
цепи, а напротив гуанина (Г) всегда находится цитозин (Ц). Комплементарное
спаривание А с Т и Г с Ц осуществляется за счет водородных связей. Между А и Т
образуется две водородные связи, между Г и Ц – три.

Комплементарность цепей ДНК составляет
химическую основу важнейшей функции ДНК – хранения и передачи генетической
информации.
Типы РНК.
Известны три основных
вида клеточных РНК: транспортные РНК (тРНК), матричные РНК (мРНК) и рибосомные
РНК (рРНК). Они различаются по месторасположению в клетке, составу и размерами,
а также функциями. РНК состоят, как правило, из одной полинуклеотидной цепи,
которая в пространстве складывается таким образом, что ее отдельные участки
становятся комплементарными друг другу (”слипаются”) и образуют короткие
двуспиральные участки молекулы, в то время как другие участки остаются
однотяжевыми.
Матричные РНК
выполняют функцию матрицы
белкового синтеза в рибосомах.
Рибосомные РНК
выполняют роль структурных
компонентов рибосом.
Транспортные РНК
участвуют в
транспортировке
a
-аминокислот из цитоплазмы в рибосомы и в переводе информации нуклеотидной
последовательности мРНК в последовательность аминокислот в белках.
Механизм передачи генетической информации.
Генетическая информация закодированиа в нуклеотидной последовательности
ДНК. Механизм передачи этой информации включает три основных этапа.
Первый этап – репликация
–копирование
материнской ДНК с образованием двух дочерних молекул ДНК, нуклеотидная
последовательность которых комплементарна последовательности материнской ДНК и
однозначно определяется ею. Репликация осуществляется путем синтеза новой
молекулы ДНК на материнской, которая играет роль матрицы. Двойная спираль
материнской ДНК раскручивается и на каждой из двух цепей происходит синтез новой
(дочерней) цепи ДНК с учетом принципа комплементарности. Процесс осуществляется
под действием фермента ДНК-полимеразы. Таким образом из одной материнской ДНК
образуются две дочерних, каждая из которых содержит в своем составе одну
материнскую и одну вновь синтезированную полинуклеотидную цепь.
Второй этап – транскрипция
– процесс, в
ходе которого часть генетической информации переписывается с ДНК в форме мРНК.
Матричная РНК синтезируется на участке деспирализованной цепи ДНК как на матрице
под действием фермента РНК-полимеразы. В полинуклеотидной цепи мРНК
рибонуклеотиды, несущие
определенные
нуклеиновые основания, выстраиваются в последовательности, определяемой
комплементарными взаимодействиями с нуклеиновыми основаниями цепи ДНК. При этом адениновому
основанию в ДНК будет соответствовать урациловое
основание в РНК. Генетическая информация о синтезе белка закодирована в ДНК с
помощью триплетного
кода. Одна аминокислота кодируется
последовательностью из трех нуклеотидов, которую называют кодоном
.
Участок ДНК, кодирующий одну полипептидную цепь, называется геном
.
Каждому кодону ДНК соответствует комплементарный кодон в мРНК. В целом молекула
мРНК комплементарна определенной части цепи ДНК – гену.
Процессы репликации и транскрипции происходят в
ядре клетки. Синтез белка осуществляется в рибосомах. Синтезированная мРНК
мигрирует из ядра в цитоплазму к рибосомам, перенося генетическую информацию к
месту синтеза белка.
Третий этап – трансляция
– процесс
реализации генетической информации, которую несет мРНК в виде последовательности
нуклеотидов в последовательность аминокислот в синтезируемом белке.
a
-Аминокислоты, необходимые для
синтеза белка транспортируются к рибосомам посредством тРНК, с которыми они
связываются путем ацилирования 3
/ -ОН группы на конце цепи тРНК.

тРНК имеет антикодоновую ветвь, содержащую
тринуклеотид — антикодон
, который соответствует переносимой ею
аминокислоте. На рибосоме тРНК прикрепляются антикодоновыми участками к
соответствующим кодонам мРНК. Специфичность стыковки кодона и антикодона
обеспечивается их комплементарностью. Между сближенными аминокислотами
образуется пептидная связь. Таким образом реализуется строго определенная
последовательность соединения аминокислот в белки, закодированная в
генах.
Нуклеотиды.
Нуклеиновые кислоты
Нуклеотиды
Нуклеотиды – это природные соединения, состоящие из 1) остатков азотистого нуклеинового основания, 2) углеводного остатка и 3) фосфатной группы.
Азотистые нуклеиновые основания
Азотистые основания – это производные двух гетероциклов - пиримидина и пурина .
Пиримидиновые основания

Пуриновые основания

ТАУТОМЕРИЯ АЗОТИСТЫХ ОСНОВАНИЙ
а) лактам-лактимная

Аналогичная таутомерия возможна у тимина, цитозина и гуанина.
б) амино-иминная

Аналогичная таутомерия возможна у гуанина и цитозина.
Лактамы более устойчивы, чем лактимы, а амины более устойчивы, чем имины. Все основания in vitro и in vivo существуют и участвуют в обмене веществ в лактамных и аминоформах.
Производные и аналоги нуклеиновых оснований применяются в медицине как лекарственные вещества противоопухолевого действия:

Нуклеозиды
Нуклеозиды – это соединения, состоящие из остатков нуклеинового основания и углевода, связанных β- N -гликозидной связью.

Реакция образования нуклеозидов in vivo идет под действием ферментов.
В кислой среде (но не в нейтральной и не в щелочной) нуклеозиды гидролизуются, распадаясь на исходные основание и углевод. Пиримидиновые нуклеозиды гидролизуются труднее, пуриновые – легче.


Номенклатура нуклеозидов
|
Основание |
Название |
|
|
2"-Дезоксиуридин |
||
|
2"-Дезокситимидин |
||
|
2"-Дезоксицитиидин |
||
|
Аденозин |
||
|
2"-Дезоксиаденозин |
||
|
Гуанозин |
||
|
2"-Дезоксигуанозин |
Нуклеотиды
Нуклеотиды – это нуклеозиды, содержащие фосфатную группу в 5"-положении (5"-фосфоридированные нуклеозиды).
Нуклеотиды образуются in vivo в результате ферментативного фосфорилирования нуклеозидов:

Нуклеотиды гидролизуются в кислой и в щелочной средах: при кислотном гидролизе образуются основание, углевод и фосфорная кислота, а щелочной гидролиз дает нуклеозид и фосфат натрия:

Номенклатура нуклеотидов
|
Основание |
Название |
|
|
Уридин-5"-монофосфат (УМФ), уридиловая кислота |
||
|
2"-Дезоксиуридин-5"-монофосфат |
||
|
Тимидин-5"-монофосфат (ТМФ), тимидиловая кислота |
||
|
2"-Дезокситимидин-5"-монофосфат |
||
|
Цитидин-5"-монофосфат (ЦМФ), цитидиловая кислота |
||
|
2"-Дезоксицитиидин-5"-монофосфат |
||
|
Аденозин-5"-монофосфат (АМФ), адениловая кислота |
||
|
2"-Дезоксиаденозин-5"-монофосфат |
||
|
Гаунозин-5"-монофосфат (ГМФ), гуаниловая кислота |
||
|
2"-Дезоксигуанозин-5"-монофосфат |
Динуклеотиды


НАД и ФАД – коферменты, участвующие в ОВ реакциях переноса водорода в организме:


Аденозинтрифосфат (АТФ)

АТФ является аккумулятором и переносчиком энергии в биохимических реакциях.
Биологические реакции АТФ
1. Фосфорилирование – перенос фосфатных групп от АТФ на другие субстраты:

2. Гидролиз с выделением энергии, используемой в синтетазных реакциях:

Нуклеиновые кислоты
Нуклеиновые кислоты – это полинуклеотиды – полимеры, состоящие из нуклеотидных остатков, связанных сахарофосфатными сложноэфирными связями.
Схема строения полинуклеотидной цепи:

Виды НК : ДНК – содержат остатки 2 -дезоксирибозы, не содержат урацила;
Д Н К
Первичная структура ДНК
Первичная структура ДНК – это определенный порядок последовательности нуклеотидов в цепи:

В первичной структуре ядерной ДНК заложен генетический код. В процессе транскрипции он "переписывается" на информационную РНК, а затем происходит трансляция: в рибосоме на матрице информационной РНК синтезируется полипептидная цепь белка. Ключ генетического кода заключается в том, что один аминокислотный остаток в синтезируемой полипептидной цепи кодируется тремя нуклеотидными остатками (триплетом) в НК, и таким образом, с помощью 4 видов нуклеотидов кодируются 20 аминокислот.
Химические свойства нуклеиновых кислот
Сложноэфирные связи, соединяющие полинуклеотидные цепи, неустойчивы в кислой и щелочной средах, и НК в этих условиях подвергаются гидролизу:

КОМПЛЕМЕНТАРНОСТЬ АЗОТИСТЫХ ОСНОВАНИЙ
Комплементарность – это соответствие формы двух сложных линий, которые подходят друг к другу "как ключ к замку".
Комплементарные пары оснований:

В паре А-Т тимин может быть (при переходе ДНК→РНК) заменен урацилом, и пара становится А-У ("взаимозаменяемость" тимина и урацила).
Биологическое значение комплементарных взаимодействий заключается в том, что они обеспечивают точность передачи информации от одной НК к другой.
Вторичная структура ДНК
Представляет собой спираль, состоящую из двух комплементарных друг другу и антипараллельных полинуклеотидных цепей ("двойная спираль"):

Биологическая роль "двойной спирали":
1) Она обеспечивает сохранность генетической информации (ядерный нуклеопротеидный комплекс "ДНК-Гистоны");
2) Обеспечивает восстановление информации при повреждении ДНК (репарация после мутаций).
Р Н К
Виды РНК : рибосомальная, информационная, транспортная.
Рибосомальная РНК (р-РНК) – структурный материал рибосом (рибосомальный нуклеопротеидный клмплекс).
Информационная (матричная) РНК (и-РНК) – промежуточный этап в процессе трансформации информации "ДНК – белок". Она синтезируется на матрице ДНК и сама служит матрицей при синтезе белка в рибосоме. и-РНК сравнительно низкомолекулярна и не имеет развитой вторичной структуры.
Транспортная РНК (т-РНК) – низкомолекулярная РНК, выполняющая следующие функции: 1) определение "своей" аминокислоты (для каждой АК существует своя т-РНК); 2) связывание с АК и транспорт её к рибосоме; 3) определение места АК в растущей полипептидной цепи.
Транспортные РНК имеют вторичную структуру "клеверного листа":

Выступающий конец ССА-3 ОН – место связывания с карбоксильной группой АК.
Триплет нуклеотидов в крайней нижней точке – кодон, комплементарный соответствующему антикодону на и-РНК.
ЛИТЕРАТУРА:
Основная
1. Тюкавкина Н.А., Зурабян С.Э., Белобородов В.Л. и др. – Органическая химия (специальный курс), кн.2 – Дрофа, М., 2008 г., с. 157-178.
2. Н.А.Тюкавкина, Ю.И.Бауков – Биоорганическая химия – ДРОФА, М., 2007 г., с. 420-444.